Structure based drug discovery facilitated by crystallography
Posted: 15 September 2017 | Dr Sheraz Gul, Dr Sophie Zimmermann | No comments yet
Drugs exert therapeutically meaningful effects if they bind with appropriate selectively and potency to their respective targets. Although many of the assay techniques employed in early-stage drug discovery provide evidence that a drug is acting upon the target under investigation, this is often indirect and the site of action and stoichiometry is unknown…

Even incisive techniques such as surface plasmon resonance (SPR) and isothermal titration calorimetry do not provide absolute evidence that the compound is interacting with the target in the desired manner. This article explains how the gap can be filled by crystallography.
Resolving the atomic structure of a protein with a compound not only confirms target binding but also provides structural information on the location of the binding site and the precise nature of the interactions between the drug’s functional groups and the amino acids in the protein which are important for binding. These results can be used to help explain the basis of effective inhibition and to improve the potency and specificity of compounds in the drug discovery workflow. Structure based drug discovery (SBDD) has become an important element of medicinal chemistry projects, but until recently has often been limited due to its low throughput and applicability for soluble targets only.
When the first x-ray crystal structures were solved in the 1950s and 60s, these results became immediately linked to physiology and drug discovery. Max Perutz and John Kendrew were able to understand sickle cell disease by solving the structure of haemoglobin1,2 and Dorothy Hodgkin, who solved the structure of insulin,3 began considering insulin redesign for treatment of diabetes as synthetic insulins became possible. But it wasn’t until the 1990s that the first crystal structures of protein-ligand complexes were submitted to the Protein Data Bank (PDB).4 The number of complex structures increased over the following years and since the Millennium protein crystallography has become a well-established technique in drug discovery.5
Obtaining protein crystals for structure determination
A drug target has to fulfil a number of requirements for crystallisation trials to be undertaken, eg, large amounts of a stable, mono-disperse protein solution at a concentration of several milligrams per millilitre must be available.6 Finding conditions for growing well-diffracting crystals is a matter of trial and error; therefore screening approaches are applied7 To speed up this process and to minimise the amount of protein required, sample robotic screens using the vapour diffusion method in 96-well format are used. Small protein drops of 100 to 200nL are mixed with equal volumes of precipitant solution and placed in vapour equilibration with a liquid reservoir containing the same precipitant.

The precipitant concentration in the drop is lower than in the reservoir because it was mixed with the protein solution. The system moves towards equilibrium by diffusion of water vapour from the drop to the reservoir with the protein and precipitant concentration in the drop increasing. If the protein molecules assemble into a periodic lattice during this process, protein crystals grow. These crystals reach a size of between 10 and several hundred µm and tend to be fragile due to a high solvent content. Even though automation has helped to streamline the process and enables screening a wider range of crystallisation conditions, growing well-diffracting crystals still remains the major bottleneck in crystallography.8
Protein–ligand complex crystals can be obtained either by mixing the ligand with the protein when setting up the crystallisation drops (co-crystallisation), or by soaking the ligand into the protein crystal. For diffraction data collection, crystals are harvested manually under a microscope from the crystallisation drop.9 Subsequently they will be cryo-protected and flash-frozen in liquid nitrogen. Diffraction data collection is usually performed at cryogenic temperatures to minimise radiation damage. Data can either be collected at a home source or at a synchrotron such as the European Synchrotron Radiation Facility (ESRF) in Grenoble or the German Electron Synchrotron (DESY) in Hamburg.
While the former might be the method of choice for initial screening of diffraction properties, the latter is preferable when it comes to high-resolution data collection due to a higher brilliance of synchrotron radiation. Synchrotron facilities offer beamlines specifically designed for protein crystallography. Crystals are rotated in a very strong, monochromatic x‑ray beam and the resulting diffraction patterns are recorded and subsequently allow calculation of the electron density in the crystal. Atomic coordinates of the protein, as well as the ligands, are then modelled into this 3D map and the resulting model allows conclusions about the interaction site and the nature of the interactions between a potential drug and its target.
Application of crystallography in drug discovery
Once a druggable target protein has been identified, it can be screened against libraries of compounds that lead to the identification of initial hits, which are the first step on the way towards new small-molecule drugs.10,11 Molecules are selected based on their potency in biological assays, but ligand-NMR, SPR, fluorescence-based thermal shift assays and mass spectrometry are also used to identify and characterise target-ligand complexes.12,13 The active compounds that are common between these techniques are subsequently optimised iteratively towards improved affinity, selectivity, ADME-Tox and PK/PD properties.
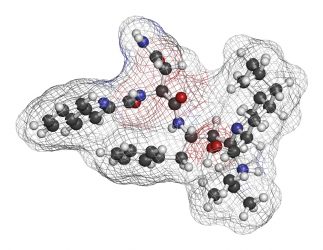
In many cases, the optimised compounds are co-crystallised with the target protein to reveal the compound-target interactions and thus offer the opportunity to improve the compound’s characteristics.14 Prominent examples of this approach include transition-state analogue inhibitors for influenza virus neuraminidase,15 adenosine-derived inhibitors of it Leishmania mexicana it glyceraldehyde-3-phosphate dehydrogenase16 and rationally designed inhibitors of HIV protease.17
The peptide HIV protease inhibitor saquinavir (Ro 31-8959, Invirase, Fortovase), from Hoffmann-La Roche, entered the clinic in 1995 and life expectancy for people with HIV and Aids began to improve immediately.18 This drug was designed based on the observation that HIV-1 protease cleaves the amino acid sequences containing dipeptides Tyr-Pro or Phe-Pro. As mammalian proteases do not cleave peptide bonds followed by a proline, this approach promised selectivity. The crystal structure of HIV-1 protease in complex with a peptide inhibitor, Ro 31-8558, suggested possible modifications. Replacement of a proline by (S,S,S)-decahydro-isoquinoline-3-carbonyl (DIQ) significantly improved the potency of the inhibitors.19 Saquinavir exhibited sub-nanomolar Ki against both HIV-1 protease and HIV-2 protease, while causing only minor inhibition of human aspartic proteases. As one of the major drawbacks associated with peptide-based drugs is proteolytic degradation, non-peptide inhibitors of HIV protease were subsequently designed by iterative co-crystal structure analysis of peptide inhibitors bound to the protease and the replacement of parts of the inhibitors by non-peptide substituents.
Fragment-based drug discovery (FBDD)
Structure determination by crystallography has become substantially faster over recent years, mainly due to developments in the fields of automation, detectors and crystallographic software. Robots can automatically image crystallisation drops, thus avoiding time-consuming manual inspection under the microscope. Moreover, there are robotic solutions under development for soaking and harvesting crystals, which can be extremely helpful, especially when dealing with very fragile membrane protein crystals. Not only have the times necessary for collecting a dataset reduced from hours to less than five minutes, there are also several synchrotrons which offer remote and even automatic data collections.
These developments have paved the way towards high-throughput approaches of crystallography and application of this method in the early screening phases of drug discovery. One such very successful screening approach is fragment-based drug discovery (FBDD).20 FBDD uses fragment libraries composed of 500-4,000 low-molecular-weight compounds (250-350Da). Crystallographic screening is performed by soaking cocktails of six to 10 fragments into apo-protein crystals. Solving the crystal structures will not only serve to identify fragment hits but simultaneously yield information about the binding site.
In 2011, the first fragment-derived drug, vemurafenib, discovered at Plexxikon and developed in partnership with Roche,21 were approved. Vemurafenib targets a mutant form of the kinase BRAF and extends life for patients with skin cancer. From an initial screen, 238 compounds were selected for co-crystal analysis and over 100 structures of kinases co-crystallised with bound compounds were determined. In the following phase iterations, co-crystallography, computational simulation, chemistry and biological assays were targeted more specifically towards the BRAF kinase system. Compounds were optimised for binding affinity, selectivity and pharmacokinetic properties, and within a year of initiating BRAF-specific improvements, vemurafenib was identified. Other examples include venetoclax and ribociclib. Venetoclax, developed by AbbVie and Genentech, binds to B-cell lymphoma 2 (BCL-2) anti-apoptotic regulators and blocks interaction with other proteins. The drug was approved by the US FDA in 2016 for the treatment of chronic lymphocytic leukaemia. Ribociclib,22 developed by Astex and Novartis, targets the protein kinase Cdk4 and will be used in combination with letrozole as a first-line treatment for advanced breast cancer.
New drug targets: membrane proteins
Membrane proteins are the gateways to the cell, controlling nutrient and waste transport across the membrane, as well as signalling cascades. Drugs frequently target membrane proteins, but the molecular interactions are often not understood.23 While the medical importance of membrane proteins has long been established, it has been impossible to crystallise these proteins because crystallisation requires purification. The first step in membrane protein purification is extraction from the lipid bilayer, which is achieved by solubilisation in detergents, but detergent belts around proteins impair crystal contact formation. Membrane protein crystallography has recently become more feasible due to several cunning methods which reduce the flexibility of these proteins and stabilise them in a specific conformation. Rather than utilising vapour diffusion approaches, membrane protein crystals are often obtained from lipid cubic phase (LCP). Here an artificial membrane system, consisting of lipid, water, and protein in appropriate proportions, forms a structured, transparent, and complex three-dimensional lipid array, which is pervaded by an intercommunicating aqueous channel system. The artificial membranes provide a matrix for the membrane proteins, allow them to cluster and support growth by lateral diffusion of protein molecules in the membrane.24,25
The 2012 Nobel Prize in Chemistry was awarded to Robert Lefkowitz and Brian Kobilka for their work on the structure and function of G-protein coupled receptors (GPCRs).26 Brian Kobilka’s group failed to crystallise the β2-adrenergic receptor-Gs protein complex in vapour diffusion but succeeded in obtaining crystals in LCP. In addition, the group also made use of a nanobody directed against the Gs protein. Nanobodies are the variable domains of heavy-chain only antibodies, found in camelids and sharks. Co-crystallising membrane proteins with these small antibody fragments has been very successful recently. The nanobody stabilises the membrane protein and also increases the hydrophilic surface, thus fostering crystal contact formation.27,28
Outlook
Membrane protein crystallography will enable the development of more specific and potent drugs for this target class and these efforts are being led by pharmaceutical companies such as Heptares Therapeutics and Receptos, who undertake SBDD for GPCRs.29 Additionally, crystallography may lead to novel routes for addressing the severe global medical problem of antibiotic resistance, as crystal structures of the ribosomal particles from Staphylococcus aureus it have revealed atomic details of drug binding sites and unique structural motifs specific for this pathogenic strain. These findings may enable the design of antibiotics specifically targeting this pathogen.30 Finally, the recent resolution revolution in single particle cryo-electron microscopy (cryo‑EM) allows the structures of very large proteins to be solved to near-atomic resolution.31 In future, this method may complement crystallography in the drug discovery process.
About the authors
Sophie Zimmermann trained as a biochemist in Hannover, Cambridge and Hamburg. She is working as a postdoc with Dr Thomas Schneider and Dr Christian Löw at the Hamburg Outstation of the European Molecular Biology Laboratory, where she develops methods for crystallographic structure determination of membrane proteins and large protein complexes.

Sheraz Gul is the Head of Drug Discovery at the Fraunhofer-IME SP, Hamburg. He has 23 years’ experience in both academia (University of London) and industry (GlaxoSmithKline). This has ranged from the detailed study of biological catalysts to the design and development of assays for high-throughput screening for the major drug target classes.
References
- Kendrew JC, Perutz MF. A comparative X-ray study of foetal and adult sheep haemoglobins. Proc R Soc Lond A Math Phys Sci. 1948 Sep 2;194(1038):375-98.
- Perutz MF, Rossmann MG, Cullis AF, Muirhead H, Will G, North AC. Structure of haemoglobin: a three-dimensional Fourier synthesis at 5.5-A. resolution, obtained by X-ray analysis. Nature. 1960 Feb 13;185(4711):416-22.
- Dodson E, Harding MM, Hodgkin DC, Rossmann MG. The crystal structure of insulin. 3. Evidence for a 2-fold axis in rhombohedral zinc insulin. J Mol Biol. 1966 Mar;16(1):227-41.
- Kemp TJ, Alcock W. 100 years of X-ray crystallography. Science Progress, Volume 100, Number 1, March 2017, pp. 25-44(20). Science Reviews 2000 Ltd.
- Thomas SE, Mendes V, Kim SY, Malhotra S, Ochoa-Montaño B, Blaszczyk M, Blundell TL. Structural Biology and the Design of New Therapeutics: From HIV and Cancer to Mycobacterial Infections: A Paper Dedicated to John Kendrew. J Mol Biol. 2017 Jun 23. pii: S0022-2836(17)30315-7. [Epub ahead of print].
- Kim Y, Babnigg G, Jedrzejczak R, Eschenfeldt WH, Li H, Maltseva N, Hatzos-Skintges C, Gu M, Makowska-Grzyska M, Wu R, An H, Chhor G, Joachimiak A. High-throughput protein purification and quality assessment for crystallization. Methods. 2011 Sep;55(1):12-28.
- Chung CW. The use of biophysical methods increases success in obtaining liganded crystal structures. Acta Crystallogr D Biol Crystallogr. 2007 Jan;63(Pt 1):62-71.
- Joachimiak A. High-throughput crystallography for structural genomics. Curr Opin Struct Biol. 2009 Oct;19(5):573-84.
- Hassell AM, An G, Bledsoe RK, Bynum JM, Carter HL 3rd, Deng SJ, Gampe RT, Grisard TE, Madauss KP, Nolte RT, Rocque WJ, Wang L, Weaver KL, Williams SP, Wisely GB, Xu R, Shewchuk LM. Crystallization of protein-ligand complexes. Acta Crystallogr D Biol Crystallogr. 2007 Jan;63(Pt 1):72-9.
- Larsen MJ, Larsen SD, Fribley A, Grembecka J, Homan K, Mapp A, Haak A, Nikolovska-Coleska Z, Stuckey JA, Sun D, Sherman DH. The role of HTS in drug discovery at the University of Michigan. Comb Chem High Throughput Screen. 2014 Mar;17(3):210-30.
- Karawajczyk A, Orrling KM, de Vlieger JS, Rijnders T, Tzalis D. The European Lead Factory: A Blueprint for Public-Private Partnerships in Early Drug Discovery. Front Med (Lausanne). 2017 Jan 19;3:75.
- Folmer RH. Integrating biophysics with HTS-driven drug discovery projects. Drug Discov Today. 2016 Mar;21(3):491-8.
- Moroni E, Paladino A, Colombo G. The Dynamics of Drug Discovery. Curr Top Med Chem. 2015;15(20):2043-55.
- Hoelder S, Clarke PA, Workman P. Discovery of small molecule cancer drugs: successes, challenges and opportunities. Mol Oncol. 2012 Apr;6(2):155-76.
- Murrell M, Porotto M, Weber T, Greengard O, Moscona A. Mutations in human parainfluenza virus type 3 hemagglutinin-neuraminidase causing increased receptor binding activity and resistance to the transition state sialic acid analog 4-GU-DANA (Zanamivir). J Virol. 2003 Jan;77(1):309-17.
- Callens M, Hannaert V. The rational design of trypanocidal drugs: selective inhibition of the glyceraldehyde-3-phosphate dehydrogenase in Trypanosomatidae. Ann Trop Med Parasitol. 1995 Dec;89 Suppl 1:23-30.
- Gu SX, Xue P, Ju XL, Zhu YY. Advances in rationally designed dual inhibitors of HIV-1 reverse transcriptase and integrase. Bioorg Med Chem. 2016 Nov 1;24(21):5007-5016.
- O’Brien WA 3rd. Saquinavir/Ritonavir: its evolution and current treatment role. AIDS Read. 2006 Jan;16(1):38-44; discussion 43.
- Ghosh AK, Kincaid JF, Walters DE, Chen Y, Chaudhuri NC, Thompson WJ, Culberson C, Fitzgerald PM, Lee HY, McKee SP, Munson PM, Duong TT, Darke PL, Zugay JA, Schleif WA, Axel MG, Lin J, Huff JR. Nonpeptidal P2 ligands for HIV protease inhibitors: structure-based design, synthesis, and biological evaluation. J Med Chem. 1996 Aug 16;39(17):3278-90.
- Malhotra S, Thomas SE(1), Ochoa Montano B(1), Blundell TL. Structure-guided, target-based drug discovery – exploiting genome information from HIV to mycobacterial infections. Postepy Biochem. 2016;62(3):262-272.
- Scheier B, Amaria R, Lewis K, Gonzalez R. Novel therapies in melanoma. Immunotherapy. 2011 Dec;3(12):1461-9.
- Deeks ED. Venetoclax: First Global Approval. Drugs. 2016 Jun;76(9):979-87.
- Molinski SV, Bozóky Z, Iram SH, Ahmadi S. Biophysical Approaches Facilitate Computational Drug Discovery for ATP-Binding Cassette Proteins. Int J Med Chem. 2017;2017:1529402.
- Landau EM, Rosenbusch JP. Lipidic cubic phases: A novel concept for the crystallization of membrane proteins. Natl. Acad. Sci. 93, 14532–14535 (1996).
- Caffrey M. A comprehensive review of the lipid cubic phase or in meso method for crystallizing membrane and soluble proteins and complexes. Acta Crystallogr. Sect. F Struct. Biol. Commun. 71, 3-18 (2015).
- Lin HH. G-protein-coupled receptors and their (Bio) chemical significance win 2012 Nobel Prize in Chemistry. Biomed J. 2013 May-Jun;36(3):118-24.
- Steyaert J, Kobilka BK. Nanobody stabilization of G protein-coupled receptor conformational states. Curr Opin Struct Biol. 2011 Aug;21(4):567-72.
- Pardon, E. et al. A general protocol for the generation of Nanobodies for structural biology. Protoc. 9, 674–693 (2014).
- Congreve M, Marshall F. The impact of GPCR structures on pharmacology and structure-based drug design. British Journal of Pharmacology. 2010;159(5):986-996.
- Khusainov I, Vicens Q, Bochler A, et al. Structure of the 70S ribosome from human pathogen Staphylococcus aureus. Nucleic Acids Research. 2016;44(21):10491-10504.
- Merk A, Bartesaghi A, Banerjee S, Falconieri V, Rao P, Davis MI, Pragani R, Boxer MB, Earl LA, Milne JLS, Subramaniam S. Breaking Cryo-EM Resolution Barriers to Facilitate Drug Discovery. Cell. 2016;165(7):1698-1707.
Related topics
Crystallography, Drug Discovery
Related organisations
European Molecular Biology Laboratory, Fraunhofer-IME SP
Related people
Dr Sophie Zimmermann, Sheraz Gul